3.2.1 Size of neuronal components
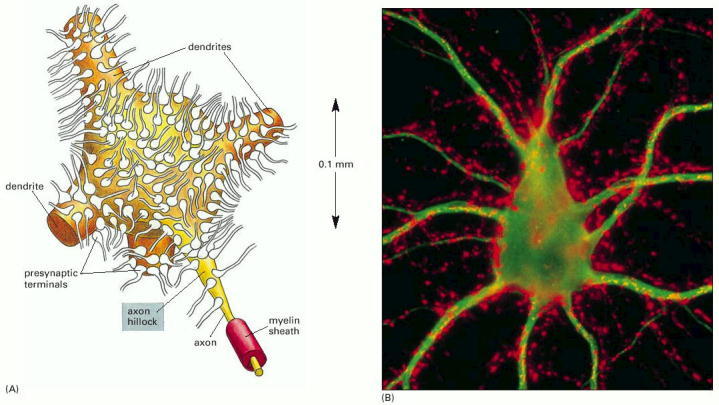
The ’dendritic trees can be quite large, containing up to 98 % of the entire neuronal surface area’ [72]. ’Because the cell body is small compared with the dendritic tree, its membrane potential is roughly uniform’ [107]; we assume that the neuron’s membrane itself is equipotential. However, the dendrites are not equipotential while delivering signals. Their potential ’is a composite of the effects of all the signals impinging on the cell, weighted according to the distances of the synapses from the cell body’. ’Temporal and spatial summation together provide the means by which … many presynaptic neurons jointly control the membrane potential.’ ’Each incoming signal is reflected in a local PSP of graded magnitude, which decreases with distance from the site of the synapse’ [107]; see our mathematical discussion of this physiological evidence in section 3.7.4. This latter sentence should read that its measurable effect (their local potential) decreases, compared to the one at the presynaptic terminals. As the surface, over which it propagates during its journey through the dendrites, extends, the charge density decreases, but the total charge is conserved until the cell body is reached. These statements mean (assuming that those signals travel with the same speed in the dendrites) that experiments underpin the presence of a ’slow current’ of ions in neurons, although the notion is not introduced.
Experimental evidence shows that the electric signals have a finite speed in axons, dendrites, and cell bodies and that within the cell, the overwhelming majority of propagation time is spent in the dendrites. The mathematical handling of finite speeds is not simple, especially within a biological cell, so we separate the cell into two regions and make the approximation that within the cell body, the interaction is instant (that is, the Laws of electricity are valid). However, outside the cell body, the finite interaction speed in the dendrites leads to observable effects that significantly influence the cell’s operation. We set up a hybrid model: the cell body is equipotential (aka: can be described by a ’fast current’), but the dendrites (and they contribute the overwhelming majority of the signal path within the cell) are non-equipotential. They must be described by approximations based on a ’slow current’. With that model, we explain the up to now not understood features of neuronal charge processing; furthermore, why is that ’the interplay between the synaptic and neuronal dynamics, when the network is near a critical point, both recurrent spontaneous and stimulated phase transitions enable the phase-dependent processes to replace each other’ [72].
The size of presynaptic terminals (reproduced here as Fig. 3.2) is about two orders of magnitude smaller than the cell body and its dendrites [107], chapter 11. In other words, the ”dendritic trees can be quite large, containing up to e98% of the entire neuronal surface area” [72]. ”Because the cell body is small compared with the dendritic tree, its membrane potential is roughly uniform” [107]. We pinpoint that since the ionic currents spend most of their travel time in the dendritic tree, we assume that the overwhelming portion of the travel time derives from the dendrites; so the contribution from the travel on the body is omitted. In this sense it is unimportant whether the membrane itself is equipotential. However, it is crucial that the dendrites are not equipotential while delivering signals. Their potential ”is a composite of the effects of all the signals impinging on the cell, weighted according to the distances of the synapses from the cell body”. ”Temporal and spatial summation together provide the means by which … many presynaptic neurons jointly control the membrane potential.” [107]. This former sentence should read that its measurable effect (their potential) decreases, compared to the one at the presynaptic terminals. As the surface, over which it propagates during its journey through the dendrites, extends, the charge density decreases, but the total charge conserves until the cell body reached. The latter sentence should read that the presynaptic terminals and the membrane potential mutually control each other. Given that the ions can reach the presynaptic terminal passively by using a ”downhill” potential between the axonal arbor and the membrane, once, they cannot enter the mebrane until the membrane’s potential is higher than that in the axonal arbor and twice, whwn they can enter, the current depends on the potential difference between the membrane and the arbor.
{kind=link}