Even at writing this text (at the midle of 202), textbooks comprise the old and bad cellular structure.
Membranes are fundamental in many places, from biological objects to industrial filters. They operate on the border of microscopic and macroscopic worlds, separating non-living and living matters, and combining electrical and thermodynamical interactions. We show that an extremely thin skin near to the surface of biological membranes is responsible for the biological thermoelectric processes.
To describe those complex and continuous phenomena at least approximately, we must separate them to stages. Using omissions, approximations and abstractions, we can describe the stages approximately, usually considering only one dominant phenomenon. The described phenomena are interrelated in a very complex way and depend on different parameters. To some point, we can describe that thin layer using a static picture and providing an empirical description of its individual processes, even we can give some limited validity mathematical descriptions for those stages. However, we understand that for describing the transition (contrasting with step-like stage changes) between those well-defined stages of the athmosphere we need a dynamic description and we need to find out the laws of motion governing the processes.
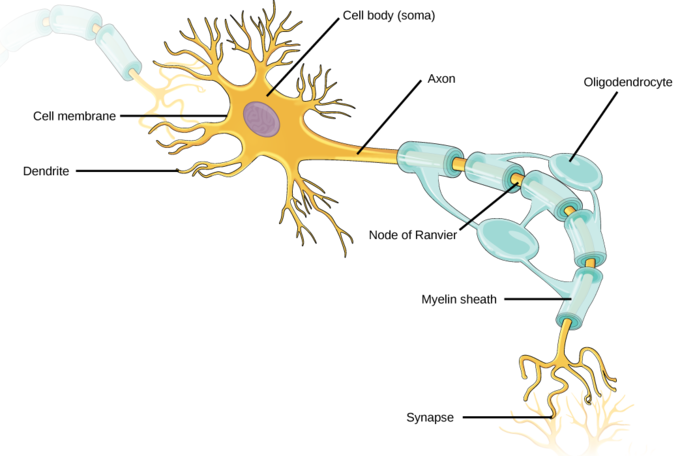
Similar is the case with the neuronal membranes and the neuronal operation. Now we are at the point where their decades-old static description is not sufficient. To descibe the neuron’s dynamic behaviour, we need to derive the corresponding laws of motion. We need a meticulous and unusual analyzis to derive them.
In a neuron, in the abstraction science uses, we put together only ionic solution, semipermeable membrane and currents reaching them. As experienced, at some combination of their parameters, qualitatively different phenomena happen, which, in the abstraction biology uses, called signs of life. Given that the approximations, the derived abstractions and the mathematical formalisms describing them are different for the two cases, it looks like we have two different, only loosely bound worlds. However, if we realize we arrived at the boundary of non-living and living matters, we must go back to the first principles of science. Using our approach, maybe we can defy that ”the emergence of life cannot be predicted by the laws of physics” [13].
The layers, for their regular operation, have both source and drain. In neurons, the source is distributed over the surface of the layers and the drain is concentrated at the terminating end of the layer. The two currents are flowing simultaneously, i.e., the source of the drain current has a time course, so the product of the two currents can be measured. (actually, it is a differential equation, and in the elementary cross-section, Kirchoff’s Junction Law is valid). Generally, it takes time until the source current reaches the drain’s position.
Initially, biology used the abstraction that the measured resistance and capacitance are distributed along the membrane’s surface. It assumed a discrete equipotential membrane with capacity and that it leaks through a discrete resistance . However, in biology, no discrete elements for storing charge exist. The notion of storing charge can be used only in the sense that for the time of passing a finite-size element with finite propagation speed, the charge carriers spend the corresponding time in the element. That phenomenon resembles storing the charge, and that imitation enables us to describe a behavior resemblant to that of the biological circuit. Attempting to imitate the effects of biological “slow” currents using electric parallels hides that generating an AP is their native feature; furthermore, slow currents may also play a role in cognitive functions.
{kind=link}