2.9.5 Currents
Resting current
Although through the thickness of the ion layer , the ion’s water-related behavior can influence the surface charge density, one can draw the conclusion that when only one type of ion is solved in the liquid, approximately the same concentration difference between the segments produces approximately the same ”thermodynamic electric field” that can counterbalance the electric field of the biological condenser. One can notice that in the classic measurement by Hodgkin-Huxley, the sum of the concentrations of and in the cytoplasm and extracellular fluid (in mM) are 380 and 390, respectively, while the sum concentrations of and are 450 and 460; practically all equal, as our analysis derived it must be. The deviations are well within the uncertainty of the measured values.
HH did a complete measurement, and derived precise measured data. However, their measurement was not accurate because their model was wrong. In a balanced state, no current flows, so no voltage is generated. For this reason, they assumed that a permanent leaking current was flowing on the membrane, and on its resistance, it generated a voltage [76]. In HH’s picture, the question what ion constitutes the current remains open. It was claimed that the only permeability pathways open at rest are and . It follows that at the resting membrane potential, the leak current equals the potassium current. That is, at rest, two currents flow (without driving force in a balanced state), one consisting of ions and another of one ions (??) (depending on the direction, it could be positive or negative), and neither of them changes the concentration neither on the departure nor on the arrival segment, neither the concentration of the other ions; defying also Nernst’s law.
The idea of leakage current was wrong: the voltage HH measured [76] is correct, but – as our model correctly explains – it was generated by charge separation instead of a current through a distributed resistor. It is permanently present while ionic segments are on its sides, even when no current flows. Measuring the energy consumption of neuronal operation [78] confirms that no resting current, in the sense as HH used the notion, exists.
Heat emission/absorption
As explained in section 2.5.2, there are alternative methods of producing a voltage difference across the membrane’s surface; with and without current. Hodgkin and Huxley took the wrong choice. The lack of resting current also solves the long-standing mystery of ”reversible heat release during the action potential”, which is inconsistent with the Hodgkin–Huxley model. In the framework of that theory, Ohmic currents flow through resistors that dissipate heat due to friction, no matter in which direction () the ion currents flow. As Hodgkin wrote [87]: ”Hill and his colleagues found [65] that an initial phase of heat liberation [of the action potential] was followed by one of heat absorption. […] a net cooling on open-circuit was totally unexpected and has so far received no satisfactory explanation.”
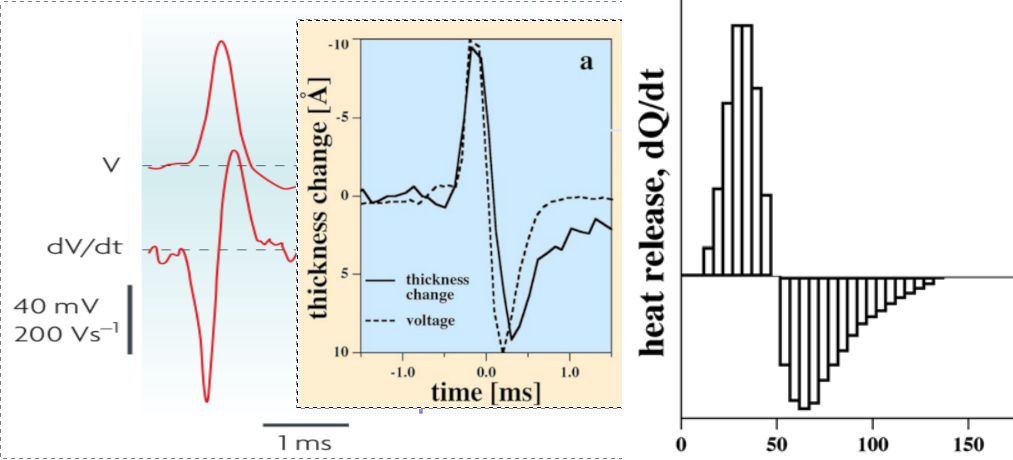
The issue here is that the dissipation was attributed to the ”leakage current.” (Although, due to the different conduction mechanisms, it is not dissipated in the way electron energy is dissipated in solids.) However, as our explanation shows, the voltage generated across the membrane is due to charging instead of a permanently flowing current, so no dissipation was present given that no resting current was present. Although the energy to build a potential on the membrane and to maintain it needs biological energy consumption, the process of generating an action potential is, in principle, reversible: ions flow into the layer on the membrane and flow out through the AIS.
According to that, the ”temperature of individual particles” with the mean kinetic energy is (where is the Boltzmann constant and is the thermodynamic temperature of the bulk quantity of the substance); that is the temperature is directly proportional to the kinetic energy. The ions on the two sides of the membrane are in the state with temperature before and after the AP. When the ions rush into the intracellular space, they gain energy through electrostatic acceleration by the membrane’s electric potential, so the same ions appear on the intracellular side as having more energy, i.e., slightly higher temperature. In the second phase of the AP, those more energetic ions that provide excess voltage above the resting potential leave the membrane through the AIS, so the temperature decreases in the second phase. The heat liberation and absorption is a direct experimental evidence of the ”slow current” and the mechanisms described above.
In the framework of the classic theory, Ohmic currents flow through resistors that dissipate heat due to friction, no matter in which direction () the ion currents flow. The existence of ”cooling”, alone, undermines the credibility of the classic theory [138]. The issue here is that the dissipation was attributed to the ”leakage current.” As Hodgkin wrote: ”Hill and his colleagues found [65] that an initial phase of heat liberation [of the action potential] was followed by one of heat absorption. […] a net cooling on open-circuit was totally unexpected and has so far received no satisfactory explanation.” [87] ”All authors came to similar conclusions: during the action potential, no significant net heat is produced. Transient heat releases are mostly reabsorbed in the second phase of the action potential. …The finding of a reversible heat release during the action potential of nerves is a striking and very fundamental fact. It is inconsistent with the HH model. The physics underlying the nervous impulse must rather be based on reversible processes”. [97]
Heat liberation and absorption are direct experimental evidence of the ”slow current” and the mechanisms described above. The lack of leakage current (see section 2.9.5) not only validates our model, but also solves the long-standing mystery of reversible heat release during the AP. Pressure changes accompany the electrical process and naturally account for the observed temperature changes. Our Speculations in Appendix 2.5.7 on the energy consumption of the nervous impulse underpins that the complex physical process comprises mostly reversible disciplinary processes: most of the energy of AP is stored as reversible elastic potential energy. Hence, its dissipation is also negligible: ”no significant net heat is produced” [97]. (Alternatively, it may be hypothesized that the positive ions, while traveling ahead of the positive ions under the effect of a potential-free space in the second phase of AP, not only turn with their positive charge the offset voltage to negative, but on the way the ions collect the heat dissipated by the ions. ”Laws of science cannot describe life”.)
The shocking conclusion in [139], that ”brain cells communicate with mechanical pulses, not electrical signals”, more precisely sounds that mechanical pulses (pressure waves) deliver the charge carriers of the electrical signals, in close interaction with the repulsion of the delivered charged carriers. In Equation (2.7), we also must consider the mechanical force delivered by the pressure wave. Furthermore, the nervous pulse transmission over the axons must be revisited. The classical mechanism based on coordinated action of ion channels is undoubtedly wrong. The enormous pressure, the incompressible electrical fluid, and the repulsion of the delivered particles suggest an alternative and more reasonable mechanism.